การอ้างอิง: ปกฉัตร ชูติวิศุทธิ์. (2563). “ปรอทและจุลินทรีย์” บทบาทของจุลินทรีย์ในการขับเคลื่อนวัฏจักรทางชีวธรณีเคมีของปรอทในสิ่งแวดล้อม. วารสารสิ่งแวดล้อม, ปีที่ 24 (ฉบับที่ 2).
บทความ: “ปรอทและจุลินทรีย์” บทบาทของจุลินทรีย์ในการขับเคลื่อนวัฏจักรทางชีวธรณีเคมีของปรอทในสิ่งแวดล้อม
ปกฉัตร ชูติวิศุทธิ์
สถาบันวิจัยสภาวะแวดล้อม จุฬาลงกรณ์มหาวิทยาลัย
E-mail: Pokchat.C@chula.ac.th
บทนำ
ปรอท คือธาตุโลหะหนักที่มีความเป็นพิษต่อสิ่งมีชีวิต และเป็นหนึ่งในสารมลพิษที่มีการกระจายตัวปนเปื้อนในสิ่งแวดล้อมจากกิจกรรมในภาคอุตสาหกรรม เช่น การทำเหมืองทอง การเผาถ่านหิน กระบวนการผลิตปูนซีเมนต์ กระบวนการผลิตเหล็กและโลหะ กระบวนการคลออัลคาไลน์เพื่อการผลิตคลอรีนและโซดาไฟ กระบวนการกลั่นน้ำมัน รวมถึงการใช้ปรอทในผลิตภัณฑ์ต่าง ๆ เช่น หลอดฟลูออเรสเซนต์ แบตเตอรี่ และวัสดุที่ใช้ในการอุดฟัน (อ้างอิงข้อมูลจาก https://www.epa.gov/international-cooperation/mercury-emissions-global-context#types) โดยปรอทในรูปใด ๆ ก็ตามสามารถก่อความเป็นพิษต่อสิ่งมีชีวิต โดยเฉพาะปรอทอินทรีย์ที่มักพบในรูปของเมทิลเมอร์คิวรี่ (Methylmercury) ซึ่งเป็นสารที่มีพิษต่อระบบประสาท (อ้างอิงจาก Committee on the Toxicological Effects of Methylmercury, 2000) และนับเป็นปรอทที่มีความเป็นพิษมากที่สุดเมื่อเทียบกับปรอทในรูปอื่น ๆ เนื่องจากมีความสามารถในการละลายในไขมันและสามารถจับกับโปรตีนในเซลล์ จึงถูกขับออกจากร่างกายได้ยากและสามารถสะสมได้เป็นปริมาณมาก ทั้งยังก่อความเป็นพิษได้ที่ความเข้มข้นซึ่งต่ำกว่าปรอทอนินทรีย์มาก (Boening, 2000; Fitzgerald and Clarkson, 1991)
อย่างไรก็ตาม เนื่องจากปรอทเป็นธาตุตามธรรมชาติที่พบได้ในเปลือกโลก การกระจายตัวของปรอทในสิ่งแวดล้อมจึงสามารถเกิดขึ้นได้จากปรากฎการณ์ทางธรรมชาติด้วยเช่นกัน เช่น จากการปะทุของภูเขาไฟ การพังทลายของดิน วัฏจักรทางอุทกวิทยา และจากไฟป่า (Dash and Das, 2012) โดยปรอทที่พบในเปลือกโลกจะมีความเข้มข้นอยู่ที่ประมาณ 21 ส่วนในพันล้านส่วน ในเปลือกโลกชั้นล่าง และประมาณ 56 ส่วนในพันล้านส่วน ในเปลือกโลกชั้นบน (Wedepohl, 1995) ซึ่งกรมควบคุมมลพิษได้กำหนดมาตรฐานคุณภาพดินที่ใช้ประโยชน์เพื่อการอยู่อาศัยและเกษตรกรรม ว่าสามารถมีปรอทและสารประกอบปรอทได้ไม่เกิน 23 ส่วนในล้านส่วน (อ้างอิงจากเว็บไซต์กรมควบคุมมลพิษ, http://www.pcd.go.th/info_serv/reg_std_soil01.html) โดยปรอทที่พบในธรรมชาติ โดยทั่วไปจะอยู่ในรูปของธาตุปรอท (Hg0) หรือในรูปของสารประกอบซัลไฟด์ประเภทซินนาบาร์ (Cinnabar) ซึ่งเป็นของแข็ง แร่ปรอทเหล่านี้มีความสามารถในการละลายน้ำต่ำ แต่ส่วนของปรอทที่ละลายออกมาได้จะสามารถอยู่ได้ทั้งในรูปประจุ +2, +1 และ 0 รวมทั้งสามารถจับกับไอออนและสารอินทรีย์อื่น ๆ ผ่านกระบวนการตามธรรมชาติหรือจากกิจกรรมของมนุษย์ (Barkay et al., 2003) และสามารถจะเคลื่อนย้ายถ่ายเทไปสู่สิ่งแวดล้อมได้ทั้งทางแหล่งน้ำ ดิน และในอากาศ
ซึ่งไอออนของปรอทที่มีประจุบวก จะสามารถจับกับไอออนอื่นที่มีประจุลบ เช่น ไฮดรอกไซด์ คลอไรด์ และซัลไฟด์ เกิดเป็นสารประกอบต่าง ๆ เช่น Hg(OH)2, HgCl2, HgSO4 และ HgS ส่วนสารประกอบปรอทอินทรีย์ มักพบในรูปของโมโนเมทิลเมอร์คิวรี่ (CH3Hg+) และไดเมทิลเมอร์คิวรี่ ((CH3)2Hg) ซึ่งสารอินทรีย์ปรอทเหล่านี้นอกจากจะมีความเป็นพิษสูง ยังมีความสามารถในการเคลื่อนที่ในสิ่งแวดล้อมและสามารถสะสมได้ในสิ่งมีชีวิต (Bioaccumulation) โดยการสะสมจะยิ่งเพิ่มปริมาณมากขึ้นในสิ่งมีชีวิตที่อยู่ด้านบนของห่วงโซ่อาหาร ผ่านการถ่ายเทสารมลพิษเป็นทอด ๆ ภายในห่วงโซ่ (Biomagnification) นอกจากนี้ ไอออนของปรอทยังสามารถจับได้กับอนุภาคและสารอินทรีย์ละลายน้ำ (Dissolved organic matter) ซึ่งการจับตัวของปรอทกับสารเหล่านี้ออกมาเป็นปรอทสปีชี่ส์ต่าง ๆ (Mercury speciation) เป็นปัจจัยที่มีผลอย่างมากต่อการหมุนเวียนวัฏจักรของปรอทในธรรมชาติ ทั้งในแง่ของปรอทที่จะถูกจับไว้ภายในระบบนิเวศและในแง่ของการขับเคลื่อนปรอทออกไปยังสิ่งแวดล้อมอื่น ๆ (Skyllberg, 2012)
โรค “มินามาตะ” บทเรียนการปนเปื้อนของปรอทจากอดีต
การปนเปื้อนของเมทิลเมอร์คิวรี่ในสิ่งแวดล้อม เคยสร้างปรากฏการณ์ก่อโรค “มินามาตะ” ในมนุษย์ โดยเหตุการณ์นี้ถูกค้นพบที่เมืองมินามาตะ ประเทศญี่ปุ่น ในปี ค.ศ.1956 หลังจากพบว่ามีผู้คนล้มป่วยเป็นจำนวนมากจากการได้รับเมทิลเมอร์คิวรี่เข้าสู่ร่างกาย ผ่านการบริโภคอาหารทะเลที่ได้จากอ่าวมินามาตะ ซึ่งมีปริมาณปรอทสะสมอยู่ในช่วง 5-35 ส่วนในล้านส่วน โดยเกิดจากการปล่อยน้ำเสียของโรงงานสารเคมีขนาดใหญ่ในพื้นที่ จนทำให้พบปริมาณปรอทสะสมในเส้นผมของผู้คนที่อาศัยในบริเวณนั้นสูงถึง 700 ส่วนในล้านส่วน (Harada, 1995) ซึ่งนับเป็นความเข้มข้นที่สูงมากเมื่อเทียบกับเส้นผมของคนทั่วไปที่จะพบปรอทได้เพียง 2 ส่วนในล้านส่วน (Hong et al., 2012) การสะสมของปรอทในสิ่งมีชีวิตก่อให้เกิดความผิดปกติในการรับความรู้สึก เกิดภาวะกล้ามเนื้อเสียสหการ มีอาการเซ มือสั่น ลิ้นแข็ง พูดไม่ชัด รวมถึงเกิดปัญหาด้านการมองเห็นและการได้ยิน เหตุการณ์นี้นำมาซึ่งการตื่นตัวต่อประเด็นการปนเปื้อนเมทิลเมอร์คิวรี่ในสิ่งแวดล้อม ซึ่งการปนเปื้อนสามารถเกิดขึ้นได้ทั้งจากการทิ้งปรอทในรูปเมทิลเมอร์คิวรี่โดยตรงผ่านกิจกรรมของมนุษย์ หรือจากการที่ปรอทอื่น ๆ ถูกเปลี่ยนรูปเป็นเมทิลเมอร์คิวรี่ผ่านกระบวนการทางชีวธรณีเคมีในวัฏจักรทางธรรมชาติ
วัฏจักรชีวธรณีเคมีของปรอท ผ่าน 4 กระบวนการหลักของจุลินทรีย์
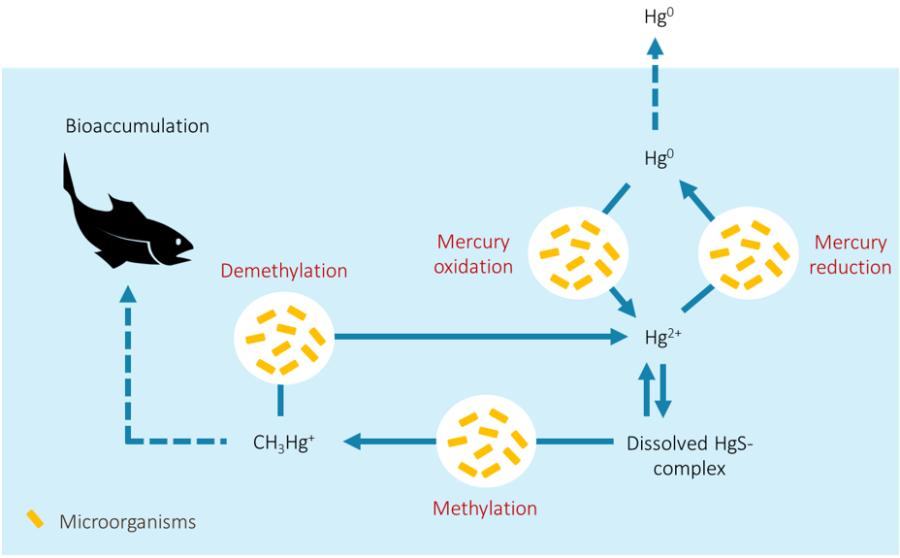
การเปลี่ยนรูปของปรอท เป็นกระบวนการที่เกิดขึ้นหมุนเวียนเป็นวัฎจักร หรือที่เรียกกันว่าวัฏจักรชีวธรณีเคมี (Biogeochemical cycle) ที่เกิดจากปฏิกิริยาทั้งทางชีววิทยา ธรณีวิทยา และทางเคมีที่มีต่อธาตุ ๆ หนึ่ง ส่งผลให้เกิดการเปลี่ยนรูปฟอร์มและเกิดการเคลื่อนที่ของธาตุนั้นในสิ่งแวดล้อม โดยวัฏจักรชีวธรณีเคมีของธาตุหลักต่าง ๆ (เช่น คาร์บอน ไนโตรเจน ซัลเฟอร์) ล้วนมีกิจกรรมทางชีววิทยาซึ่งขับเคลื่อนโดยสิ่งมีชีวิตขนาดเล็กจำพวกจุลินทรีย์เข้ามาเกี่ยวข้อง และนับเป็นสิ่งมีชีวิตหลักที่มีบทบาทในการหมุนเวียนธาตุ ให้เกิดการเปลี่ยนรูปผ่านการเปลี่ยนสถานะรีดอกซ์ (Redox state) และการเปลี่ยนรูประหว่างสารอินทรีย์และสารอนินทรีย์ การเปลี่ยนรูปนี้ทำให้คุณสมบัติทางฟิสิกส์และเคมีของธาตุนั้น ๆ เปลี่ยนแปลงไป ก่อให้เกิดการเคลื่อนที่ของธาตุระหว่างตัวกลางในธรรมชาติ อันได้แก่ ดิน น้ำ อากาศ และสิ่งมีชีวิต แม้แต่โลหะหนักที่มีความเป็นพิษอย่างปรอท ก็มีการขับเคลื่อนวัฏจักรชีวธรณีเคมีผ่านทางจุลินทรีย์เช่นเดียวกัน โดยกระบวนการหลักที่มีจุลินทรีย์เข้ามาเกี่ยวข้องในการเปลี่ยนรูปปรอท แสดงไว้ดังรูปที่ 1
รูปที่ 1 วัฏจักรชีวธรณีเคมีของปรอท ผ่านการขับเคลื่อนโดยกระบวนการต่าง ๆ ของจุลินทรีย์
กระบวนการทางชีวธรณีเคมีของปรอทที่ขับเคลื่อนโดยจุลินทรีย์ สามารถออกแบ่งได้เป็น 4 กระบวนการหลัก ได้แก่ 1) กระบวนการเปลี่ยนรูปไอออนของปรอท (Hg2+) เป็นรูปปรอทไม่มีประจุ (Hg0) หรือเมอร์คิวรี่รีดักชัน (Mercury reduction) 2) กระบวนการเปลี่ยนรูป Hg2+ เป็นเมทิลเมอร์คิวรี่ หรือเมทิลเลชัน (Methylation) 3) กระบวนการเปลี่ยนรูปเมทิลเมอร์คิวรี่เป็น Hg2+ หรือดีเมทิลเลชัน (Demethylation) และ 4) กระบวนการเปลี่ยนรูป Hg0 เป็น Hg2+ หรือเมอร์คิวรี่ออกซิเดชัน (Mercury oxidation) โดยกระบวนการเหล่านี้ต่างถูกขับเคลื่อนโดยวิถีเมแทบอลิซึม (Metabolic pathway) เฉพาะ กล่าวคือแต่ละกระบวนการมีเอนไซม์ซึ่งใช้ในการเร่งปฏิกิริยาที่แตกต่างกัน
กระบวนการเมอร์คิวรี่รีดักชัน
ในพื้นที่ที่มีการสะสมตัวของปรอท มักพบจุลินทรีย์ที่สามารถเร่งปฏิกิริยาเมอร์คิวรี่รีดักชัน ผ่านกลไกการป้องกันตัวเองจากพิษปรอทของจุลินทรีย์ ซึ่งขับเคลื่อนโดยกลุ่มยีน mer ที่ประกอบไปด้วยยีนหลักคือ merA ที่มีหน้าที่ในการสร้างเอนไซม์ Mercuric reductase ซึ่งจะเปลี่ยน Hg2+ ที่มีความไวต่อการเกิดปฏิกิริยาและมีความสามารถในการละลายน้ำสูง ไปเป็น Hg0 อะตอมเดี่ยวที่มีความเสถียรต่อการเกิดปฏิกิริยามากกว่า ทั้งมีความสามารถในการละลายน้ำต่ำและมีความดันไอสูง จึงเกิดการแพร่ออกจากเซลล์ได้ง่าย (Barkay et al., 2003) ปรอทในรูปนี้จึงมักหลุดจากแหล่งน้ำหรือตะกอนขึ้นไปสู่อากาศ และเป็นรูปของปรอทหลักที่พบเจอได้ในชั้นบรรยากาศ โดยพบว่าปฏิกิริยาทางชีวภาพที่เปลี่ยน Hg2+ ไปเป็น Hg0 นี้เป็นกระบวนการสำคัญที่ทำให้เกิดการเคลื่อนที่ของปรอทระหว่างตัวกลางต่าง ๆ (Skyllberg, 2012)
ทั้งนี้ ปรอทในรูป Hg0 ยังสามารถถ่ายเทไปยังส่วนอื่น ๆ ของโลกได้ทางอากาศ และกลับลงสู่พื้นดินได้ผ่านทางฝนและหิมะ ซึ่งเป็นเส้นทางหลักในการกระจายตัวของปรอทบนพื้นผิวโลก (Lalonde et al., 2002) โดยส่วนใหญ่ปรอทในฝนและหิมะจะอยู่ในรูปของ Hg2+ นอกจากนี้ จุลินทรีย์ที่อาศัยอยู่ในชั้นบรรยากาศ (เช่น จุลินทรีย์ที่อาศัยอยู่ในระบบนิเวศของก้อนเมฆ ซึ่งมีความเข้มข้นอยู่ที่ประมาณ 102 - 105 เซลล์ต่อมิลลิลิตร (Amato, 2012)) ยังสามารถทำให้เกิดปฏิกิริยาเมอร์คิวรี่รีดักชัน ทำให้ปรอท Hg2+ กลับมาอยู่ในรูป Hg0 จึงเป็นอีกหนึ่งวัฏจักรที่หมุนเวียนปรอทในชั้นบรรยากาศ และป้องกันไม่ให้ปรอทส่วนหนึ่งตกกลับลงสู่พื้นผิวโลก
นอกจากยีน merA แล้ว ในจุลินทรีย์ที่สามารถปกป้องตัวเองจากปรอทได้เหล่านี้ยังมียีนอื่น ๆ ในกลุ่มยีนmer ได้แก่ ยีน merT, merC, merP, merH, merF และ merE ซึ่งเป็นยีนที่มีหน้าที่สร้างโปรตีนสำหรับนำปรอทเข้าสู่เซลล์ของจุลินทรีย์ และมียีน merR เป็นยีนที่คอยควบคุมการทำงานของยีนต่าง ๆ ในกลุ่ม mer รวมถึงยีน merB ซึ่งมีบทบาทสำคัญในการสร้างเอนไซม์ที่สามารถเปลี่ยนปรอทในรูปเมทิลเมอร์คิวรี่ให้กลับมาอยู่ในรูป Hg2+ ที่สามารถจะถูกเปลี่ยนรูปต่อให้เป็น Hg0 ได้โดยยีน merA (Barkay et al., 2003; Barkay et al., 2010; Dash and Das, 2012; Skyllberg, 2012)
จุลินทรีย์ที่มีกลไกต้านทานปรอทด้วยการเปลี่ยนรูปปรอทให้มีความเสถียรมากขึ้นนี้ เป็นจุลินทรีย์ที่พบเจอได้ทั้งในจุดที่เกิดการปนเปื้อนปรอทจากกิจกรรมของมนุษย์ รวมถึงพื้นที่ที่เกิดปรอทจากกิจกรรมความร้อนใต้พื้นผิวโลก เช่น ใต้ทะเลลึกและในน้ำพุร้อน อย่างไรก็ตาม พบว่าจุลินทรีย์ที่มียีน merA ส่วนใหญ่จัดอยู่ในกลุ่มที่ใช้ออกซิเจนในการหายใจ และใช้สารอินทรีย์คาร์บอนในการดำรงชีวิต (Aerobic heterotrophic microorganism) โดยมักไม่พบยีนนี้ในจุลินทรีย์ที่ดำรงชีวิตแบบไร้อากาศ (Anaerobic microorganism) ซึ่ง Barkay et al. (2010) ได้ทำการวิเคราะห์ยีน merA ที่มีอยู่บนฐานข้อมูลจีโนมและลำดับนิวคลีโอไทด์ พบยีน merA ในจุลินทรีย์จำพวกอาร์เคีย (Archaea) ในไฟลัม Euryarcheota และ Crenarcheota โดยพบในออร์เดอร์ Thermoplasmatales และ Sulfolobales ตามลำดับ ซึ่งทั้งสองออร์เดอร์ของอาร์เคียนี้มักพบในระบบนิเวศน้ำพุร้อนที่มีความเป็นกรด (Dworkin et al., 2006) และเป็นกลุ่มอาร์เคียที่ใช้ออกซิเจนในการหายใจเป็นส่วนใหญ่
ในส่วนของแบคทีเรีย พบ merA ในไฟลัม Proteobacteria ทั้งในคลาส Alpha, Beta และ Gamma รวมถึงในไฟลัม Firmicutes และ Actinobacteria ซึ่งล้วนเป็นไฟลัมหลักของแบคทีเรีย รวมทั้งพบยีน merA ในไฟลัม Deinococcus/Thermus, Aquificae, Bacteriodetes, Chloroflexi, Nitrospirae และ Verrucomicrobia โดย Barkay et al. (2010) ได้ให้คำอธิบายไว้ว่ายีน merA นั้นจำกัดวงอยู่ในจุลินทรีย์ที่สามารถดำรงชีวิตภายใต้สภาวะแวดล้อมที่ปรอทจะอยู่ในรูป Hg2+ ซึ่งเป็นรูปของปรอทที่มีความไวต่อการเกิดปฏิกิริยา ส่งผลให้จุลินทรีย์ที่มีวิวัฒนาการในการต้านทานปรอท เป็นกลุ่มที่อาศัยอยู่ภายใต้สภาวะทางรีดอกซ์และค่าพีเอชที่ขับเคลื่อนให้ปรอทอยู่ในรูป Hg2+ เป็นหลัก ในขณะที่สภาวะแวดล้อมซึ่งขับเคลื่อนให้ปรอทอยู่ในรูปที่เสถียรกว่า เช่น Hg0 หรือเมอร์คิวรี่ซัลไฟด์ (HgS) มักไม่พบจุลินทรีย์ที่มียีน merA จึงกล่าวได้ว่ารูปฟอร์มของปรอทเป็นปัจจัยหลักในการคัดเลือกจุลินทรีย์ที่มีความสามารถในการดำรงอยู่ภายใต้สภาวะนั้น ๆ
กระบวนการเมทิลเลชัน
นอกจากยีนกลุ่ม mer แล้ว ยีนที่นักวิจัยซึ่งศึกษากระบวนการชีวธรณีเคมีของปรอทให้ความสนใจกันมาก คือกลุ่มยีนที่สามารถเปลี่ยนรูปของปรอทให้เป็นเมทิลเมอร์คิวรี่ได้ ซึ่งก็คือยีน hgcA และ hgcB โดยยีนทั้งสองตัวนี้ทำหน้าที่ร่วมกันในการสร้างเมทิลเมอร์คิวรี่จาก Hg2+ ภายในเซลล์ของจุลินทรีย์ โดยพบว่าจุลินทรีย์กลุ่มหลักในสิ่งแวดล้อมที่มีบทบาทในการสร้างเมทิลเมอร์คิวรี่อยู่ในกลุ่มแบคทีเรียที่ใช้ซัลเฟตในการหายใจ หรือที่เรียกว่าปฏิกิริยารีดักชัน (Sulfate-reducing bacteria) (Barkay et al., 2003; Skyllberg, 2012)
นอกจากนี้ ยังมีการค้นพบว่าแบคทีเรียที่ใช้ไอออนของเหล็กในการหายใจ (Iron-reducing bacteria) และจุลินทรีย์สร้างมีเทน (Methanogen) ก็มีบทบาทในการสร้างเมทิลเมอร์คิวรี่ในสิ่งแวดล้อมเช่นกัน (Bravo et al., 2018) โดยกลุ่มแบคทีเรียที่ใช้ซัลเฟตในการหายใจและสามารถสร้างเมทิลเมอร์คิวรี่ได้นั้นพบว่าอยู่ในคลาส Deltaproteobacteria ทั้งหมด (Widdel and Bak, 1992) อย่างไรก็ตาม การสร้างเมทิลเมอร์คิวรี่ไม่ใช่คุณสมบัติที่พบได้ในแบคทีเรียทุกตัวที่ใช้ซัลเฟตในการหายใจ (Benoit et al., 2003) เพียงแต่เป็นคุณสมบัติที่พบในแบคทีเรียหลายสปีชี่ส์ภายในกลุ่มนี้
ทั้งจุลินทรีย์ที่ใช้ซัลเฟต ไอออนของเหล็ก และกลุ่มสร้างมีเทน ล้วนเป็นจุลินทรีย์ที่เติบโตภายใต้สภาวะไร้อากาศ โดยในงานวิจัยที่ผ่านมาพบว่า รูปฟอร์มของปรอทในสภาวะไร้อากาศจะถูกควบคุมโดยปฏิกิริยาระหว่างไอออนของปรอทกับซัลไฟด์และสารอินทรีย์ธรรมชาติ (Natural organic matter) เป็นหลัก ซึ่งปฏิกิริยาเหล่านี้จะเป็นตัวกำหนดปริมาณปรอทที่ใช้ได้ในทางชีวภาพ (Bioavailability) โดยปรอทในรูป Hg2+ มีความสามารถสูงมากในการจับกับสารประกอบซัลไฟด์และธาตุหมู่ 7 (Halides) (Skyllberg, 2012) ส่วนในสารอินทรีย์ธรรมชาติ ปรอทจะสามารถจับกับหมู่ซัลฟ์ไฮดริล (Sulfhydryl functional groups) ซึ่งมีซัลเฟอร์เป็นองค์ประกอบ (Hsu-Kim et al., 2013)
โดยปฏิกิริยาเหล่านี้จะส่งผลให้เกิดสารประกอบปรอททั้งที่อยู่ในรูปละลายน้ำ ในรูปอนุภาคระดับนาโน (Nanoparticulate) และในรูปผลึกอนุภาค (Crystalline particle) ซึ่งการเปลี่ยนรูปฟอร์มของปรอทระหว่างสารประกอบต่าง ๆ และระหว่างสถานะละลายน้ำกับสถานะของแข็ง คือปัจจัยที่ส่งผลโดยตรงต่อปริมาณปรอทที่จุลินทรีย์จะนำไปใช้ได้ในระบบนิเวศนั้น ๆ กล่าวคือจุลินทรีย์จะสามารถใช้สารประกอบปรอทในรูปที่สามารถนำเข้าสู่เซลล์ได้ผ่านกลไกต่าง ๆ ของผนังเซลล์ และมักจะเป็นรูปฟอร์มที่ละลายในน้ำ นอกจากนี้ในงานวิจัยที่ผ่านมายังพบว่า อัตราการเกิดเมทิลเมอร์คิวรี่ในน้ำและในตะกอนไม่ได้ขึ้นอยู่กับความเข้มข้นของปรอทเสมอไป เนื่องจากมีการตั้งสมมุติฐานว่ามีปริมาณปรอทเพียงเล็กน้อยเท่านั้นที่อยู่ในรูปซึ่งจุลินทรีย์กลุ่มสร้างเมทิลเมอร์คิวรี่จะสามารถนำไปใช้ได้
อย่างไรก็ตาม รูปฟอร์มของปรอทที่จุลินทรีย์สามารถใช้ได้ภายใต้สภาวะไร้อากาศนั้น ยังไม่เป็นที่เข้าใจกันมากนัก มีการตั้งทฤษฎีว่าจุลินทรีย์สร้างเมทิลเมอร์คิวรี่สามารถนำปรอทเข้าสู่เซลล์ได้โดยการลำเลียงแบบไม่ใช้พลังงาน (Passive transport) ทำให้รูปฟอร์มหลักของปรอทที่สามารถเข้าสู่เซลล์ได้ด้วยวิธีนี้ คือสารประกอบปรอทขนาดเล็กที่ไม่มีประจุ เช่น Hg(HS)20 และ HgS0 ที่ละลายน้ำ (Hsu-Kim et al., 2013) รวมถึงทฤษฎีที่กล่าวว่าจุลินทรีย์สร้างเมทิลเมอร์คิวรี่ สามารถนำปรอทเข้าสู่เซลล์ด้วยการลำเลียงแบบใช้พลังงาน (Active transport) ซึ่งปรอทที่สามารถนำเข้าสู่เซลล์ได้ด้วยวิธีนี้ ได้แก่ปรอทที่จับกับหมู่ไทออล (Thiol group) ซึ่งเป็นสารประกอบอินทรีย์ซัลเฟอร์ที่ถูกสร้างได้โดยจุลินทรีย์ และสามารถสะสมเป็นปริมาณมากภายใต้สภาวะที่มีออกซิเจนต่ำหรือไร้ออกซิเจน (Adediran et al., 2019; Skyllberg, 2012)
โดยรูปของปรอทที่พบว่าจุลินทรีย์สามารถนำไปใช้ได้ดี คือปรอทที่จับกับไทออลซึ่งมีมวลขนาดเล็ก (Low molecular mass thiol) ในขณะที่อัตราการเกิดเมทิลเมอร์คิวรี่จะลดลงเมื่อปรอทอยู่ในรูปผลึกอนุภาคซัลไฟด์ หรือจับอยู่กับหมู่ไทออลของสารอินทรีย์ขนาดใหญ่ (Manceau et al., 2015) รูปฟอร์มของปรอทและปัจจัยที่ควบคุมการนำปรอทเข้าสู่เซลล์ จึงเป็นกุญแจสำคัญที่ควบคุมการเกิดเมทิลเมอร์คิวรี่ภายใต้ระบบนิเวศแบบไร้อากาศ
จากการวิเคราะห์ฐานข้อมูลจีโนมและลำดับนิวคลีโอไทด์โดย McDaniel et al. (2019) พบว่าจุลินทรีย์สร้างเมทิลเมอร์คิวรี่ที่มีอยู่ในฐานข้อมูลปัจจุบัน ครึ่งหนึ่งจัดอยู่ในไฟลัม Deltaproteobacteria, Firmicutes, Euryarchaeota และ Bacteroidetes และโดยส่วนใหญ่เป็นแบคทีเรียที่ใช้ซัลเฟตในการหายใจ (ในคลาส Delta-proteobacteria) นอกจากนี้ยังมีจุลินทรีย์กลุ่มอื่น ๆ ซึ่งมีการค้นพบไม่นานมานี้ว่ามีบทบาทต่อการสร้างเมทิลเมอร์คิวรี่ในสิ่งแวดล้อมเช่นกัน ได้แก่ แบคทีเรียในไฟลัม Spirochaetes, Planctomycetes, Verrucomicrobia, Chlamydiae และ Lentisphaerae รวมถึงการค้นพบยีน hgcA ในแบคทีเรียกลุ่ม Nitrospirae, Chloroflexi, Elusimicrobia และ Actinobacteria ซึ่งไม่เคยมีการรายงานมาก่อนว่ามีจุลินทรีย์สร้างเมทิลเมอร์คิวรี่อยู่ภายในไฟลัม ดังนั้นจะเห็นได้ว่าการสร้างเมทิลเมอร์คิวรี่เป็นคุณสมบัติที่พบได้ในจุลินทรีย์หลากหลายชนิด นอกเหนือไปจากกลุ่มที่เป็นที่รู้จักกันดีอย่างแบคทีเรียที่ใช้ซัลเฟตในการหายใจ
กระบวนการดีเมทิลเลชัน
ปริมาณของเมทิลเมอร์คิวรี่ที่ตกค้างในสิ่งแวดล้อม เป็นผลระหว่างการทำงานของจุลินทรีย์สร้างเมทิลเมอร์คิวรี่กับจุลินทรีย์ที่สามารถทำให้เกิดดีเมทิลเลชัน หรือปฏิกิริยาที่เปลี่ยนเมทิลเมอร์คิวรี่กลับเป็น Hg2+ ซึ่งเกิดจากการทำงานของเอนไซม์ Organomercury lyase ที่สร้างขึ้นโดยยีน merB โดยยีนนี้จะพบได้ในแบคทีเรียไฟลัม Actinobacteria, Firmicutes และ Proteobacteria (Barkay et al., 2010) การทำงานของยีน merB จะช่วยลดการสะสมของเมทิลเมอร์คิวรี่ที่ถูกสร้างขึ้นในระบบนิเวศ อย่างไรก็ตาม ข้อมูลของยีน merB ในฐานข้อมูลจีโนมและลำดับนิวคลีโอไทด์ยังมีไม่มากนักเมื่อเทียบกับยีนอื่น ๆ ส่งผลให้องค์ความรู้เกี่ยวกับจุลินทรีย์ที่สามารถทำให้เกิดปฏิกิริยาดีเมทิลเลชันได้ยังมีอยู่จำกัดในปัจจุบัน
กระบวนการเมอร์คิวรี่ออกซิเดชัน
นอกจากกระบวนการต่าง ๆ ที่กล่าวมาข้างต้น ยังมีปฏิกิริยาเปลี่ยนรูปปรอทที่ขับเคลื่อนโดยจุลินทรีย์อีกกระบวนการหนึ่ง ซึ่งสามารถเปลี่ยนรูปปรอท Hg0 กลับเป็น Hg2+ ได้ หรือที่เรียกว่าปฏิกิริยาเมอร์คิวรี่ออกซิเดชัน โดยเป็นปฏิกิริยาที่มีการศึกษาน้อยที่สุดเมื่อเทียบกับกระบวนการเปลี่ยนรูปปรอทอื่น ๆ แต่นับเป็นปฏิกิริยาที่มีความสำคัญในแง่ของการเพิ่มปริมาณ Hg2+ และลดปริมาณ Hg0 ในระบบนิเวศ ซึ่ง Hg2+ จากปฏิกิริยานี้จะสามารถเวียนกลับเข้าสู่กระบวนการเมอร์คิวรี่รีดักชันหรือกระบวนการเมทิลเลชันได้ ทั้งนี้ขึ้นอยู่กับสภาวะแวดล้อมที่ Hg2+ อยู่ในขณะนั้น โดยเมอร์คิวรี่ออกซิเดชันเกิดจากการทำงานของเอนไซม์ Hydroperoxidases ที่สร้างขึ้นโดยยีน katG และ katE ซึ่งจากการศึกษาของ Smith et al. (1998) พบว่าแบคทีเรียที่พบได้ในดินที่อากาศเข้าถึงอย่าง Streptomyces และ Bacillus มีปฏิกิริยาการเปลี่ยน Hg0 เป็น Hg2+ ในอัตราที่สูง แสดงให้เห็นถึงความสามารถในการเกิดเมอร์คิวรี่ออกซิเดชันในดิน อย่างไรก็ตาม พบว่าในจุลินทรีย์ที่มีกลไกต้านทานปรอท จะเกิดอัตราเมอร์คิวรี่ออกซิเดชันต่ำกว่าเมอร์คิวรี่รีดักชันอย่างน้อย 10 เท่า (Barkay et al., 2003) เมื่ออ้างอิงจากข้อมูลที่มีอยู่ในปัจจุบันจะเห็นได้ว่า โอกาสในการเปลี่ยน Hg0 เป็น Hg2+ จะเกิดขึ้นได้น้อยกว่าปฏิกิริยาจาก Hg2+ เป็น Hg0
กระบวนการทางชีวธรณีเคมีอื่น ๆ
นอกจากปฏิกิริยาทางชีวภาพแล้ว การเปลี่ยนรูปของปรอทยังสามารถเกิดขึ้นได้ผ่านปฏิกิริยารีดักชันแบบใช้แสง (Photoreduction) โดยพบว่าภายใต้สภาวะแวดล้อมที่มีแสงสว่างส่องถึง แสงจะเป็นปัจจัยหลักที่ขับเคลื่อนการเปลี่ยนรูปของปรอท ซึ่งสามารถจะถูกเปลี่ยนรูปจาก Hg2+ เป็น Hg0 ได้เช่นเดียวกับปฏิกิริยาของจุลินทรีย์ รวมถึงการเปลี่ยนรูป Hg0 เป็น Hg2+ ก็สามารถเกิดผ่านปฏิกิริยาออกซิเดชันแบบใช้แสง (Photooxidation) ได้เช่นกัน (Vost et al., 2012) แสงจึงเป็นอีกหนึ่งปัจจัยสำคัญที่ส่งผลต่อรูปฟอร์มและการเคลื่อนที่ของปรอทในสิ่งแวดล้อมนั้น ๆ นอกเหนือไปจากปฏิกิริยาที่เกิดขึ้นทางเคมีและทางชีวภาพ
จากวัฏจักรชีวธรณีเคมีของปรอทที่ขับเคลื่อนโดยจุลินทรีย์ซึ่งกล่าวมาข้างต้น จะเห็นได้ว่ามีกลุ่มจุลินทรีย์หลากหลายชนิดที่มีบทบาทในการเปลี่ยนรูปและการเคลื่อนที่ของปรอทระหว่างตัวกลางต่าง ๆ ทั้งภายในระบบนิเวศที่มีและไม่มีออกซิเจน รวมถึงการเปลี่ยนรูปของปรอททางเคมีภายใต้สภาวะหนึ่ง ๆ ก็เป็นอีกปัจจัยสำคัญที่ส่งผลต่อการเกิดรูปฟอร์มของปรอทที่จุลินทรีย์จะสามารถนำไปใช้ได้ ซึ่งปฏิกิริยาเหล่านี้มีการเกิดขึ้นและเปลี่ยนแปลงอยู่ตลอดเวลาภายในระบบนิเวศ และเป็นพลศาสตร์สำคัญที่ขับเคลื่อนวัฏจักรของปรอทภายในโลกของเรา การจะแก้ปัญหาการปนเปื้อนของปรอทในสิ่งแวดล้อมจึงจำเป็นต้องมีความเข้าใจต่อกลไกทางธรรมชาติเหล่านี้ เพื่อให้รู้ถึงรูปของปรอทที่เกิดขึ้นและเส้นทางที่ปรอทในรูปฟอร์มต่าง ๆ จะไปได้ เพื่อให้สามารถวางแผนการจัดการรวมถึงการบำบัดพื้นที่ปนเปื้อนปรอทได้อย่างมีประสิทธิภาพ
เอกสารอ้างอิง
Adediran, G.A., Liem-Nguyen, V., Song, Y., Schaefer, J.K., Skyllberg, U. and Bjorn, E. (2019). Microbial biosynthesis of thiol compounds: implications for speciation, cellular uptake, and methylation of Hg(II). Environmental Science & Technology, 53, 8187−8196.
Amato, P. (2012). Clouds provide atmospheric oases for microbes. Microbe, 7(3), 119−123.
Barkay, T., Miller, S.M. and Summers, A.O. (2003). Bacterial mercury resistance from atoms to ecosystems. FEMS Microbiology Reviews, 27, 355–384.
Barkay, T., Kritee, K., Boyd, E. and Geesey, G. (2010). A thermophilic bacterial origin and subsequent constraints by redox, light and salinity on the evolution of the microbial mercuric reductase. Environmental Microbiology, 12(11), 2904–2917.
Benoit, J.M., Gilmour, C.C., Heyes, A., Mason, R.P. and Miller, C.L. (2003) Geochemical and biological controls over methylmercury production and degradation in aquatic ecosystems. In: Chai, Y. and Braids, O.C., editors, Biogeochemistry of Environmentally Important Trace Elements, 262–297. Washington: American Chemical Society.
Boening, D.W. (2000). Ecological effects, transport, and fate of mercury: a general review. Chemosphere, 40, 1335–1351.
Bravo, A.G., Peura, S., Buck, M., Ahmed, O., Mateos-Rivera, A., Ortega, S.H., Schaefer, J.K., Bouchet, S., Tolu, J., Björn, E. and Bertilsson, S. (2018). Methanogens and iron-reducing bacteria: the overlooked members of mercury-methylating microbial communities in boreal lakes. Applied and Environmental Microbiology, 84(23), e01774-18.
Dash, H.R. and Das, S. (2012). Bioremediation of mercury and the importance of bacterial mer genes. International Biodeterioration & Biodegradation, 75, 207–213.
Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, C.H. and Stackebrandt, E. (2006). The Prokaryotes. Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes. New York, USA: Springer New York.
Fitzgerald, W.F. and Clarkson, T.W. (1991). Mercury and monomethylmercury: present and future concerns. Environmental Health Perspectives, 96, 159–166.
Harada, M. (1995). Minamata disease: methylmercury poisoning in Japan caused by environmental pollution. Critical Reviews in Toxicology, 25(1), 1–24.
Hong, Y., Kim, Y. and Lee, K. (2012). Methylmercury exposure and health effects. Journal of Preventive Medicine and Public Health, 45(6), 353–363.
Hsu-Kim, H., Kucharzyk, K.H., Zhang, T. and Deshusses, M.A. (2013) Mechanisms regulating mercury bioavailability for methylating microorganisms in the aquatic environment: a critical review. Environmental Science & Technology, 47, 2441−2456.
Lalonde, J.D., Poulain, A.J. and Amyot, M. (2002). The role of mercury redox reactions in snow on snow-to-air mercury transfer. Environmental Science & Technology, 36, 174–178.
Manceau, A., Lemouchi, C., Enescu, M., Gaillot, A.-C., Lanson, M., Magnin, V., Glatzel, P., Poulin, B.A., Ryan, J.N., Aiken, G.R., Gautier-Luneau, I. and K.L. Nagy. (2015). Formation of mercury sulfide from Hg(II)−thiolate complexes in natural organic matter. Environmental Science & Technology, 49, 9787−9796.
McDaniel, E.A., Peterson, B., Stevens, S.L.R., Tran, P.Q., Anantharaman, K. and McMahon, K.D. (2019). Expanded phylogenetic diversity and metabolic flexibility of microbial mercury methylation. doi: https://doi.org/10.1101/2020.01.16.909358.
Skyllberg, U. (2012). Chemical speciation of mercury in soil and sediment. In: Liu, G., Cai, Y. and O’Driscoll, N. editors, Environmental chemistry and toxicology of mercury (2nd), 219–258. Hoboken, New Jersey: John Wiley & Sons, Inc.
Smith, T., Pitts, K., McGarvey, J.A. and Summers, A.O. (1998). Bacterial oxidation of mercury metal vapor, Hg(0). Applied and Environmental Microbiology, 64, 1328−1332.
Vost, E.E., Amyot, M. and O’Driscoll, N.J. (2012). Photoreactions of mercury in aquatic systems. In: Liu, G., Cai, Y. and O’Driscoll, N. editors, Environmental chemistry and toxicology of mercury (2nd), 219–258. Hoboken, New Jersey: John Wiley & Sons, Inc.
Wedepohl, K.K. (1995). The composition of the continental crust. Geochimica et Cosmochimica Acta, 59, 1217–1232.
Widdel, F. and Bak, F. (1992). Gram-negative mesophilic sulfate-reducing bacteria. In: Balows, A., Truper, H.G., Dowrkin, M., Harder, W. and Schleifer, K.H. editors, The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications (2nd), 3352–3378. New York: Springer-Verlag.





